文献分享|多组学揭示香蕉皮褐变过程中低温诱导的 WRKY 因子调控网络重构
文章题为"Low temperature-induced regulatory network rewiring via WRKY regulators during banana peel browning"。该文章发表于 Plant Physiology(中科院一区,IF=6.5,第一作者:Wenjun Zhu,通讯作者: Peitao Lü)。
文献内容简介
香蕉作为典型的热带水果,对低温十分敏感。低温不仅能够破坏细胞结构,还会导致严重的褐变现象。目前,热带水果对低温的反应机制与模式植物的冷响应机制之间的差异尚不明确。本研究旨在深入探究香蕉在低温环境下的分子响应机制。
研究发现,在低温条件下,香蕉皮会遭受细胞结构的破坏和显著的褐变,这主要与细胞内环境的失衡和氧化应激有关。通过多组学分析,作者揭示了香蕉皮在低温诱导下染色质可及性的改变、组蛋白修饰的模式、远端顺式调控元件的活性、转录因子的结合特性以及基因表达水平的显著波动。特别值得注意的是,WRKY 转录因子家族在这一过程中发挥了关键作用。它们通过与冷诱导基因的启动子和增强子区域结合,调控了与褐变相关的基因表达。此外,本研究还建立了香蕉在低温条件下的全基因组 DNase I 高敏感位点图谱,并构建了调控网络。这些成果不仅为理解热带植物对冷胁迫的响应提供了新的视角,而且为提高热带水果的冷耐受性和延长其货架寿命提供了重要的分子基础。
本研究作者综合分析了室温和低温条件下香蕉果皮的生理变化、转录组学、组蛋白修饰和染色质可及性,以深入探讨低温储藏对香蕉果皮褐变的影响及其分子调控机制。实验观察到,在低温条件下,香蕉果皮亮度降低,LOX 和 PPO 活性显著增加,H2O2 水平和 MDA 含量上升,这些生理指标的变化与果皮褐变现象密切相关。通过 RNA-seq 分析,作者发现多个与冷应激和褐变相关的基因在低温响应中的表达水平显著增加,特别是一些已知的冷响应基因,如 MaCBF1、MaCBF2、MaCBF3、MaPPO1、MaPOD2 和 MaLOX6。差异表达基因分析显示,在低温条件下,大量基因的表达水平发生显著变化,其中许多基因与褐变途径相关,并显示出时间上的特异性响应。k-means 聚类分析将差异表达基因(DEGs)分为六组,其中下调基因主要涉及植物发育和能量代谢,而上调基因则与磷脂降解、活性氧产生和酶促褐变等过程密切相关。超过一半的褐变相关基因在上调聚类中,表明这些基因在低温诱导的果皮褐变中起着关键作用。
ChIP-seq 分析揭示了低温条件下香蕉果皮中染色质结构的变化,活性组蛋白标记如 H3K4me3、H3K9ac 和 H3K27ac 在基因的启动子区域高度富集,而抑制性组蛋白标记 H3K27me3 则主要分布在基因体中。ChromHMM 软件定义了 8 种不同的染色质状态,将基因组中的基因分为活性、抑制和无信号三类。全局分析 DNase I 高敏感位点的分布情况进一步揭示了低温条件下香蕉果皮中染色质可及性的动态变化及其与基因表达的关系。大约 70%的 DHSs 集中在转录起始位点(TSS)附近,而近 20%位于基因间隔区域。在低温处理过程中,TSS 周围的染色质可及性显著增加,表明低温迅速改变了香蕉果皮中的染色质状态。尽管超过 75%的 DHSs 与 H3K27ac 和 H3K9ac 修饰位点重叠,但近一半的 DHSs 并未被这两种活性组蛋白修饰覆盖,暗示 H3K27ac 和 H3K9ac 可能不是香蕉果皮中活性增强子的特异性标志。结合 ChromHMM 软件定义的染色质状态,冷诱导的 DHSs 的聚类分析发现,这些 DHSs 在 TSS 周围的活性染色质区域表现出显著变化,并且与至少一种活性组蛋白修饰标记相关,这些区域的基因表达水平也相应提高。
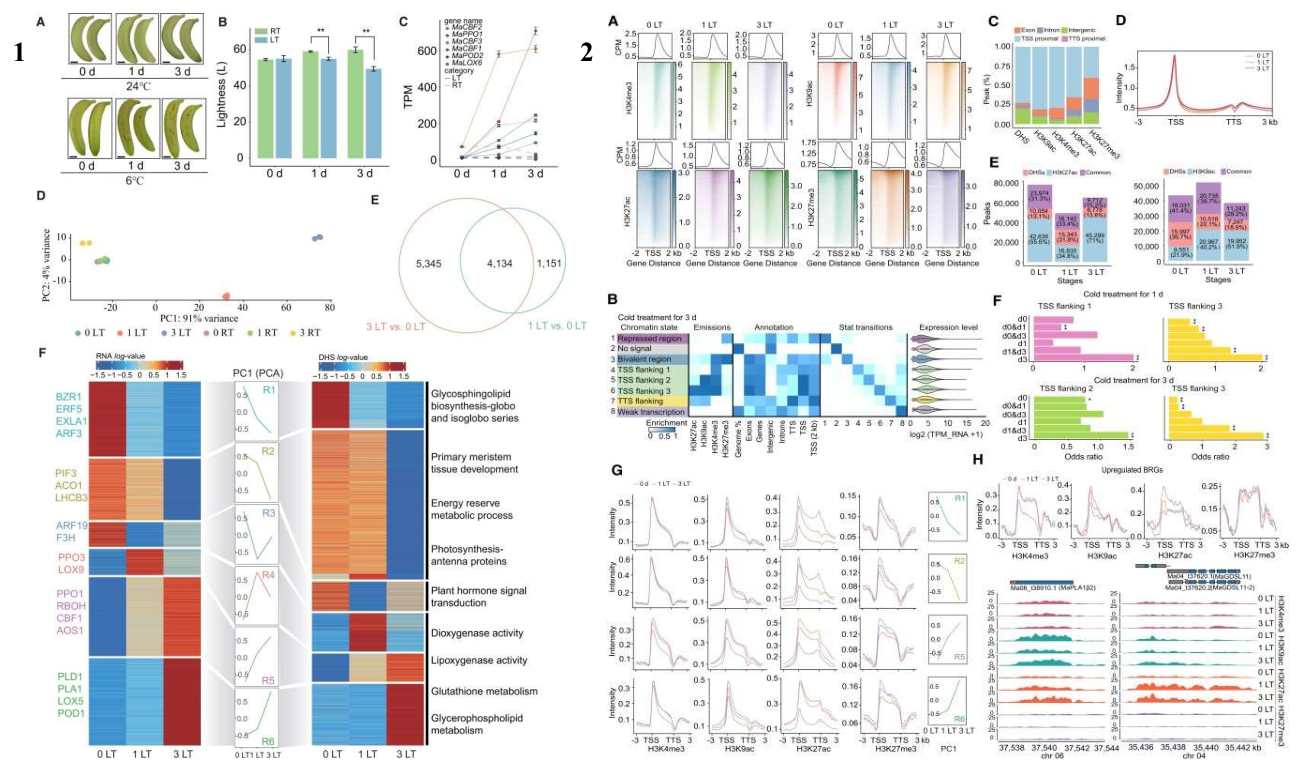
作者通过综合分析差异表达基因(DEGs)的表观遗传特征,深入探讨了低温条件下香蕉果皮中染色质可及性和组蛋白修饰的变化。主成分分析将 DNase I 高敏感位点(DHSs)分为六个模块,发现与冷响应相关的模块在冷暴露后显示出染色质可及性的增加。在 5 号和 6 号聚类中,上调的 DEGs 表现出 H3K4me3、H3K9ac 和 H3K27ac 水平的增加,这与基因的高转录水平一致。相反,下调的 DEGs 在转录起始位点(TSS)周围显示出 H3K9ac 和 H3K27ac 水平的降低。此外,研究还发现许多 DHSs 位于启动子区域之外,可能作为增强子发挥作用。这一点通过活性接触模型分析 Hi-C 和 DNase-seq 数据集得到了验证。进一步的瞬时表达实验在普通烟草叶片中验证了这些候选增强子的转录增强子活性。研究还鉴定了冷诱导的和时间特异性的增强子,并发现这些增强子与褐变相关基因的启动子区域结合,参与了 BRGs 表达的调控。例如,MaGDE1 和 MaPLA1II5 在低温暴露后受到远端增强子的调控,其表达水平增加。
作者发现,染色质的可接近区域在香蕉基因组中广泛分布,为转录因子提供了结合热点。使用 chromVAR 软件,作者识别出低温响应的基序活性。结果表明,在冷处理过程中,ARF、AGL 和 GATA 转录因子的结合位点减少,而 WRKYs、NACs、CBFs、TCPs、PCFs 和 ERFs 等转录因子的结合位点在 3 天的冷处理后富集。这些转录因子家族在多种植物对冷响应中扮演核心调控角色。基因组足迹分析进一步揭示了低温下香蕉果皮中顺式调控元件的功能变化。TOBIAS 框架的应用显示,从 0 天时的 ATHB、AGL、GRF 和 ARF 转录因子结合到它们相应的基序,转变为在 6°C 下 1 天后 WRKY 和 ERF 转录因子的结合,表明低温促进了 WRKY 和 ERF 家族成员的结合和可能的激活。TOBIAS 映射还显示,在上调的聚类中,WRKY 和 NAC 转录因子的结合在冷暴露后显著富集。而在下调的聚类中,与发育相关的转录因子如 AGLs、ARFs 和 GRFs 的结合被 ERF、MYB、NAC 和 WRKY 成员在冷反应中的染色质结合所取代。RNA-seq 数据进一步支持了 WRKY 基因在低温下的特异性诱导。在低温组中,100 个 WRKY 基因差异表达,其中 38 个与常温组共享,且大多数在常温和低温组中显示出相反的表达模式。与温带水果苹果相比,香蕉果皮中的 WRKY 基因被特异性诱导,并显示出更高的表达水平。这些结果表明,WRKY 家族在香蕉对低温的反应中发挥中心作用,并与果皮褐变强烈相关。
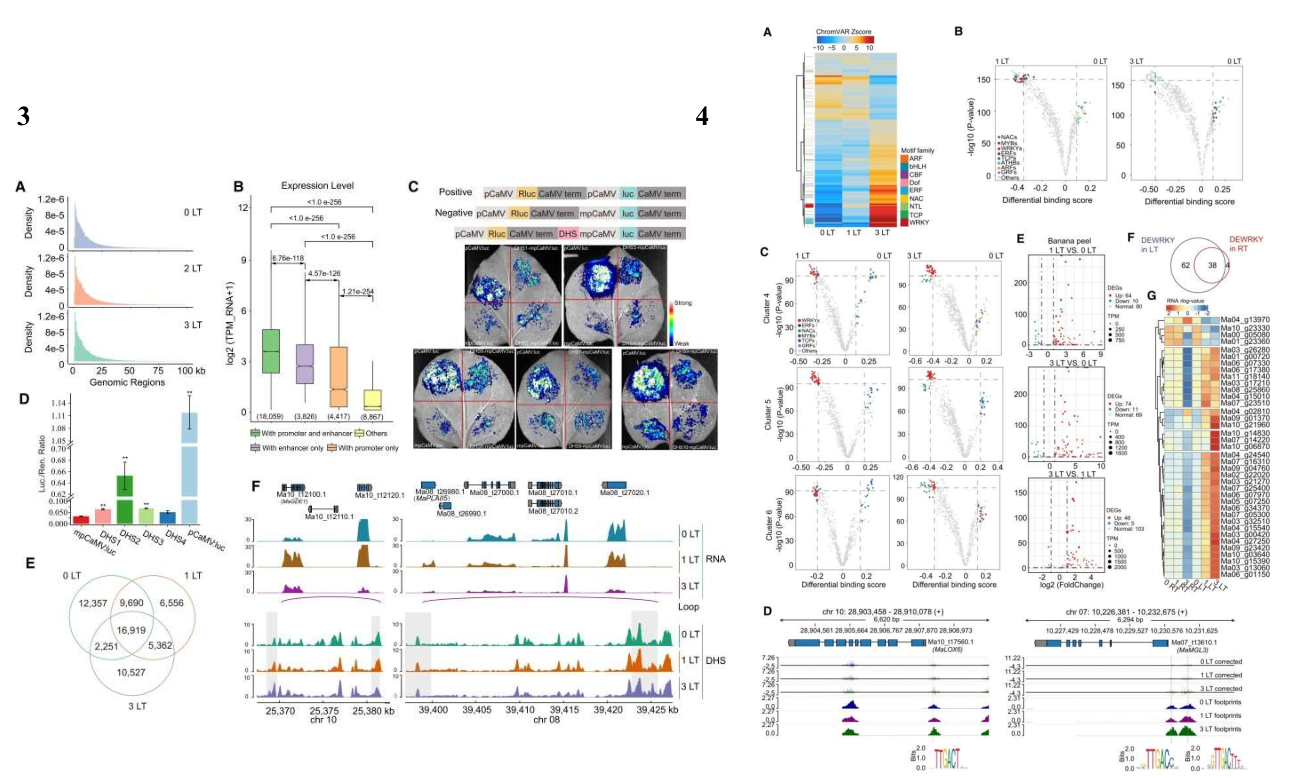
作者综合分析了 WRKY 转录因子在香蕉果皮中的调控作用,揭示了它们在提高冷耐受性和调节膜脂降解中的关键角色。通过 DAP-seq 分析,发现 MaWRKY18、MaWRKY40、MaWRKY60、MaWRKY11 和 MaWRKY33 等 WRKY 转录因子在低温下迅速上调,表达量显著增加。这些转录因子的结合位点主要集中在转录起始位点附近,且显著富含核心基序(-TTGAC-)。通过整合 RNA-seq、ChIP-seq、DAP-seq 和 DNase-seq 数据集,作者观察到 WRKY 转录因子结合到含有激活性组蛋白标记的 MaCBF1 启动子上,可能直接或间接调控 MaCBF1 在低温下的表达。同时,WRKY 转录因子也直接结合到冷诱导的 MaC2H2 启动子区域,调控其表达。在果皮褐变方面,WRKY 转录因子靶向 46 个褐变相关基因(BRGs)的启动子,并可能通过增强子-启动子相互作用,增强这些基因在低温下的表达。酵母双杂交、双分子荧光互补和 Co-IP 实验验证了 WRKY 转录因子之间的物理相互作用,表明它们可能形成复合体共同调控下游靶基因的表达。
过表达 WRKY 转录因子的实验结果表明,MaWRKYs 通过增强 BRGs 的表达促进了低温诱导的香蕉果皮褐变。相反,WRKY 转录因子的瞬时沉默减少了果皮褐变和 BRGs 的表达,进一步证实了 WRKY 转录因子在香蕉果皮褐变中的关键调控作用。通过构建以 WRKY 转录因子为核心的调控网络,作者深入探究了它们在香蕉果皮对低温响应中的协调作用。通过比较控制条件与低温处理条件下的转录因子-WRKY 互作,揭示了低温响应下转录因子网络的动态变化,分析结果发现,在低温条件下,一些原有的调控关系消失,同时形成了新的调控关系。例如,在常温下,MYB3R5 仅调控 WRKY 基因,而 WRKY 转录因子则调控与植物发育相关的 GATA12、AGL6 和 MYB61 等基因。在低温条件下,ERF4 和 HSFB2A 等转录因子参与了 WRKY 基因的诱导表达,并进一步调控其他基因。进一步分析整合了 5 个 WRKY 转录因子的 DAP-seq 数据、基因组足迹和候选增强子,详细剖析了由转录因子调控的转录因子和褐变相关基因(BRGs)的冷诱导网络。结果揭示了 10 种调控模式和 580 个调控互作,例如 WRKY18 直接调控 MaLOX5B,而 WRKY60 通过诱导 ERF019 间接控制 MaAAO3。值得注意的是,WRKY 转录因子在低温下的调控关系发生了全面的重构,与 RNA-seq 数据一致,表明 WRKY 转录因子直接调控的脂肪酶基因在低温响应中与 WRKY 基因表达谱紧密相关。此外,WRKY 转录因子直接调控的 BRGs,如 MaPLA1β2 和 MaGGDE1,也受到 WRKY 靶向增强子的调控,其表达水平在低温下增加。
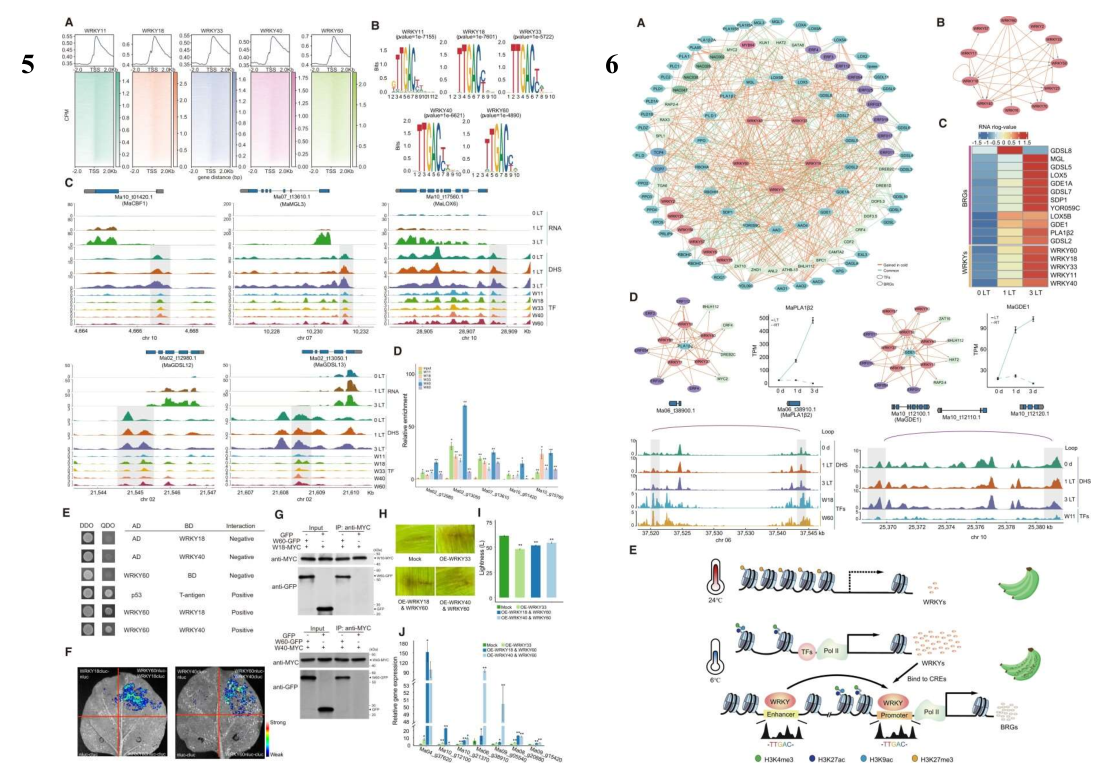
讨论总结
- 短期低温处理可诱导香蕉果皮中大量基因的表达,特别是那些与磷脂代谢和氧化有关的基因。
- 冷诱导基因通常伴随着染色质可及性的增加,以及启动子区域活性组蛋白标记(如 H3K4me3、H3K9ac 和 H3K27ac)的富集。
- WRKY 转录因子可能通过结合到褐变相关基因的启动子和增强子区域,形成染色质环,从而增强 BRGs 的表达。
- 研究发现,组蛋白 H3 的乙酰化(H3K9ac 和 H3K27ac)主要分布在转录起始位点附近,与基因的高表达水平相关,但与增强子的标记无直接联系。
- 低温改变了香蕉果皮的染色质开放性景观,特别是在基因间隔区和启动子区域,这表明远端调控元件在响应低温中发挥着关键作用。此外,低温还影响了染色质可及性景观,以及 WRKY 和 ERF 转录因子的结合模式。
- WRKY 转录因子在不同植物间的低温响应中表现出保守性,但在香蕉果皮中可能特异性地参与 CBF 表达和冷响应调节。香蕉可能利用 WRKY 转录因子从免疫反应转向应对冷胁迫的机制,但这种机制可能仅限于提高冷耐受性。如果冷耐受性失衡,可能导致细胞内稳态的紊乱,最终引起褐变。